
Blood, Heart, and Vessels: How Your Circulatory System Works
Imagine a vast, intricate network of rivers and tributaries, constantly flowing, delivering essential supplies, removing waste, and maintaining the delicate balance of an entire ecosystem. Now, shrink that image down and place it within the human body. This is the essence of our circulatory system—a masterpiece of biological engineering centered around three fundamental components: the life-giving fluid called blood, the powerful, rhythmic pump known as the heart, and the extensive network of vessels that form the circulation pathways. Together, they orchestrate the continuous movement that sustains every cell, tissue, and organ, making life itself possible. This exploration delves deep into the structure, function, and profound interconnectedness of blood, the heart, and circulation, revealing the remarkable symphony playing within us every moment of every day.
The Vital Fluid: Unveiling the Mysteries of Blood
Blood is far more than a simple red liquid. It is a complex, living tissue, a specialized connective tissue suspended in a fluid matrix, performing a staggering array of functions critical for survival. Comprising approximately seven to eight percent of an adult’s body weight (roughly five to six liters), blood is a dynamic transport medium, a communication network, a defense force, and a temperature regulator all rolled into one.
The composition of blood can be broadly divided into two main components: plasma and formed elements. Plasma, making up about fifty-five percent of the total blood volume, is the liquid extracellular matrix. It is a pale yellow fluid, primarily composed of water (about ninety percent), which serves as the solvent for a diverse array of dissolved substances. These include plasma proteins (albumins, globulins, fibrinogen), electrolytes (sodium, potassium, calcium, chloride, bicarbonate), nutrients (glucose, amino acids, lipids), waste products (urea, creatinine, bilirubin), respiratory gases (oxygen, carbon dioxide), and hormones. Albumins, the most abundant plasma proteins, are crucial for maintaining osmotic pressure, preventing excessive fluid from leaking out of capillaries into tissues. Globulins include antibodies (immunoglobulins) vital for immune defense and transport proteins (e.g., transferrin for iron). Fibrinogen is essential for the clotting process. Electrolytes are indispensable for nerve conduction, muscle contraction, maintaining pH balance, and regulating fluid distribution between compartments.

The remaining forty-five percent of blood volume consists of the formed elements, which are the cells and cell fragments suspended in the plasma. These include red blood cells (erythrocytes), white blood cells (leukocytes), and platelets (thrombocytes). Red blood cells are by far the most numerous, numbering in the trillions. Their primary, almost exclusive, function is the transport of respiratory gases. Each erythrocyte is a small, biconcave disc, a shape that provides a large surface area-to-volume ratio for efficient gas exchange and allows flexibility to squeeze through narrow capillaries. Mature red blood cells in mammals lack a nucleus and most organelles, maximizing space for their key cargo: hemoglobin. Hemoglobin is a complex iron-containing protein made up of four polypeptide chains, each bound to a heme group that holds an iron atom. It is this iron that reversibly binds to oxygen in the lungs and releases it in the tissues. The binding of oxygen to hemoglobin in the lungs (forming oxyhemoglobin) is facilitated by high oxygen partial pressure and a slightly alkaline pH. In the tissues, where oxygen partial pressure is lower and pH is slightly more acidic (due to carbon dioxide), oxygen dissociates from hemoglobin (deoxyhemoglobin). Hemoglobin also transports a portion of carbon dioxide (about twenty percent) from tissues back to the lungs, binding to it as carbaminohemoglobin. The production of red blood cells, a process called erythropoiesis, occurs primarily in the red bone marrow of adults. It is stimulated by the hormone erythropoietin (EPO), mainly produced by the kidneys in response to low blood oxygen levels. The average lifespan of a red blood cell is about one hundred twenty days, after which old and damaged cells are removed by macrophages, primarily in the spleen and liver.
White blood cells, or leukocytes, are the cellular defenders of the body. They are far less numerous than red blood cells but are crucial for immunity and inflammation. Unlike erythrocytes, leukocytes possess a nucleus and other organelles and are capable of independent movement (diapedesis) out of the bloodstream into tissues. They are broadly categorized into two main groups based on the presence or absence of granules in their cytoplasm: granulocytes and agranulocytes. Granulocytes include neutrophils, eosinophils, and basophils. Neutrophils are the most abundant white blood cells and are the first responders to bacterial infections. They are highly phagocytic, engulfing and destroying pathogens. Eosinophils combat parasitic infections and are involved in allergic reactions. Basophils release histamine (promoting inflammation) and heparin (an anticoagulant). Agranulocytes include lymphocytes and monocytes. Lymphocytes are the cornerstone of the adaptive immune system, comprising B cells (which produce antibodies), T cells (which coordinate cell-mediated immunity and kill infected cells), and natural killer (NK) cells (which destroy virus-infected cells and tumor cells). Monocytes are the largest white blood cells; they circulate in the blood for a short time before migrating into tissues, where they differentiate into macrophages or dendritic cells. Macrophages are potent phagocytes that engulf pathogens, dead cells, and debris, and also play key roles in antigen presentation and immune activation. Dendritic cells are specialized antigen-presenting cells that initiate adaptive immune responses. The production of white blood cells, leukopoiesis, also occurs in the red bone marrow and is regulated by various cytokines (colony-stimulating factors) in response to infections, inflammation, or other immune challenges. Their lifespan varies greatly, from hours to years, depending on the cell type.
Platelets, or thrombocytes, are not true cells but rather small, anucleated cell fragments derived from large bone marrow cells called megakaryocytes. They play an indispensable role in hemostasis, the process of stopping bleeding. When a blood vessel is injured, platelets adhere to the exposed collagen fibers at the injury site, become activated, change shape, and release granules containing substances like ADP, serotonin, and thromboxane A2. These chemicals attract more platelets to the site (platelet aggregation) and cause vasoconstriction, narrowing the vessel to reduce blood flow. The aggregated platelets form a temporary platelet plug. Simultaneously, the coagulation cascade is initiated, a complex series of reactions involving clotting factors (most synthesized in the liver) present in the plasma. This cascade culminates in the conversion of soluble fibrinogen into insoluble fibrin threads. These threads form a mesh that traps blood cells, platelets, and plasma, forming a stable clot that seals the break in the vessel wall. Once the vessel is repaired, fibrinolysis, the breakdown of the clot, occurs, restoring normal blood flow. Platelets typically survive for about ten days in circulation.
Beyond its core functions of transport (oxygen, carbon dioxide, nutrients, hormones, waste), defense (immune cells, antibodies, clotting), and regulation (pH balance via buffers, temperature via distribution of heat), blood also plays roles in fluid balance and communication. It truly is the river of life, connecting every part of the body and maintaining the internal environment necessary for cellular function.
The Tireless Pump: Exploring the Marvel of the Heart
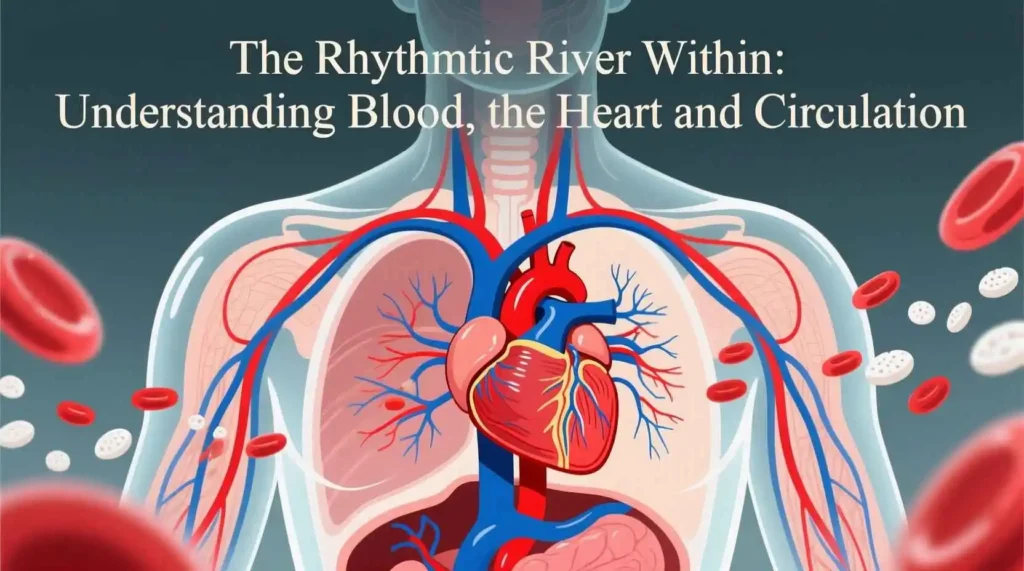
At the center of the circulatory system lies the heart, a remarkable muscular organ about the size of a closed fist, weighing roughly 250 to 350 grams. Positioned slightly left of the midline in the thoracic cavity, between the lungs and above the diaphragm, the heart functions as a dual pump, tirelessly contracting and relaxing to propel blood throughout the entire circulatory network. Its rhythmic beating, initiated before birth and continuing ceaselessly until death, is the defining sound and pulse of life.
The heart is enclosed within a protective double-walled sac called the pericardium. The outer fibrous pericardium is a tough, dense connective tissue layer that anchors the heart within the mediastinum and prevents overfilling. The inner serous pericardium consists of two layers: the parietal layer, which lines the inner surface of the fibrous pericardium, and the visceral layer (also known as the epicardium), which adheres tightly to the heart muscle itself. Between these two serous layers is the pericardial cavity, containing a small amount of lubricating serous fluid (pericardial fluid) that reduces friction as the heart beats.
The wall of the heart itself consists of three distinct layers: the epicardium, the myocardium, and the endocardium. The epicardium, as mentioned, is the visceral layer of the serous pericardium. It is composed of a thin layer of mesothelial cells and underlying connective tissue, often containing fat. The myocardium is the middle layer and by far the thickest. It is composed of specialized cardiac muscle tissue (cardiac muscle cells or cardiomyocytes) responsible for the heart’s powerful contractions. Cardiac muscle cells are striated like skeletal muscle but are branched and interconnected by specialized junctions called intercalated discs. These discs contain desmosomes (which hold cells together) and gap junctions (which allow rapid passage of ions and electrical impulses, enabling the heart to contract as a single coordinated unit, a functional syncytium). The myocardium’s thickness varies depending on the workload of the specific chamber; it is thickest in the left ventricle, which pumps blood to the entire systemic circulation. The innermost layer is the endocardium, a smooth, thin layer of endothelial cells (simple squamous epithelium) overlying a thin layer of connective tissue. This smooth lining is continuous with the endothelium of the blood vessels, minimizing friction as blood flows through the heart chambers.
Internally, the heart is divided into four chambers: two upper atria (singular: atrium) and two lower ventricles. The atria are thin-walled chambers that primarily receive blood returning to the heart. The right atrium receives deoxygenated blood from the systemic circulation via two large veins: the superior vena cava (draining the head, neck, and upper limbs) and the inferior vena cava (draining the trunk and lower limbs). It also receives blood from the heart muscle itself via the coronary sinus. The left atrium receives oxygenated blood from the lungs via four pulmonary veins (usually two from each lung). The ventricles are the thick-walled, powerful pumping chambers that eject blood out of the heart. The right ventricle pumps deoxygenated blood to the lungs, while the left ventricle pumps oxygenated blood to the systemic circulation. The internal structure of the ventricles includes raised muscular ridges called trabeculae carneae and cone-shaped papillary muscles. The papillary muscles project into the ventricular cavities and are connected to the edges of the atrioventricular valves by chordae tendineae, strong fibrous cords.
Valves within the heart ensure the one-way flow of blood, preventing backflow. There are two types of valves: atrioventricular (AV) valves and semilunar (SL) valves. The AV valves are located between the atria and ventricles. The right AV valve, also called the tricuspid valve, has three cusps (flaps). The left AV valve, also called the bicuspid or mitral valve, has two cusps. When the ventricles are relaxed (diastole), the AV valves are open, allowing blood to flow from the atria into the ventricles. When the ventricles contract (systole), the pressure within them rises, forcing the AV valve cusps closed. The chordae tendineae and papillary muscles prevent the valve cusps from everting (prolapsing) back into the atria during ventricular contraction. The semilunar valves are located at the bases of the large arteries leaving the ventricles. The pulmonary semilunar valve is at the exit of the right ventricle into the pulmonary trunk. The aortic semilunar valve is at the exit of the left ventricle into the aorta. These valves consist of three pocket-like cusps. When the ventricles contract, pressure forces the SL valves open, allowing blood to be ejected into the pulmonary trunk and aorta. When the ventricles relax, blood starts to flow back toward the heart, filling the cusps of the SL valves and causing them to snap shut, preventing backflow into the ventricles. The characteristic “lub-dup” sounds of the heartbeat are caused by the closing of these valves: the “lub” (first heart sound, S1) is the closure of the AV valves, and the “dup” (second heart sound, S2) is the closure of the semilunar valves.
The heart’s ability to beat rhythmically and automatically is due to its intrinsic conduction system, a specialized network of non-contractile cardiac muscle cells that initiate and distribute electrical impulses throughout the myocardium. This system ensures the coordinated contraction of the atria first, followed by the ventricles, maximizing pumping efficiency. The key components are:
1. Sinoatrial (SA) Node: Located in the wall of the right atrium near the entrance of the superior vena cava, the SA node is the heart’s natural pacemaker. It spontaneously generates electrical impulses at a regular rate (typically 60-100 times per minute at rest), setting the basic rhythm of the heart.
2. Atrioventricular (AV) Node: Located in the inferior part of the interatrial septum, near the opening of the coronary sinus. The impulse from the SA node spreads across the atria via gap junctions in the myocardium, causing atrial contraction. The impulse then reaches the AV node. The AV node delays the impulse for about 0.1 seconds. This delay is crucial as it allows the atria to fully contract and empty their blood into the ventricles before the ventricles begin to contract.
3. Atrioventricular (AV) Bundle (Bundle of His): From the AV node, the impulse enters the AV bundle, the only electrical connection between the atria and ventricles. It is located at the top of the interventricular septum.
4. Right and Left Bundle Branches: The AV bundle divides into the right and left bundle branches, which extend down the interventricular septum towards the apex of the heart. The left bundle branch further divides into anterior and posterior fascicles.
5. Purkinje Fibers: The bundle branches terminate in a vast network of Purkinje fibers that spread throughout the ventricular myocardium, starting from the apex and moving upwards. These fibers rapidly conduct the impulse, causing the ventricular muscle cells to contract almost simultaneously from the apex upwards, efficiently ejecting blood towards the semilunar valves.
This sequence of electrical activation generates an electrical current that spreads through the body and can be detected and recorded by an electrocardiogram (ECG or EKG). The ECG provides a graphical representation of the heart’s electrical activity, showing distinct waves: the P wave (atrial depolarization), the QRS complex (ventricular depolarization), and the T wave (ventricular repolarization). The ECG is a vital diagnostic tool for assessing heart rhythm, detecting conduction abnormalities, and identifying signs of ischemia or infarction.
The heart’s pumping action is described by the cardiac cycle, a sequence of events encompassing one complete heartbeat, including both contraction (systole) and relaxation (diastole). The cardiac cycle consists of several phases:
1. Late Diastole (Ventricular Filling): Both atria and ventricles are relaxed. Blood flows passively from the veins into the atria and then through the open AV valves into the ventricles. About seventy to eighty percent of ventricular filling occurs passively during this phase.
2. Atrial Systole: The SA node fires, the atria contract, forcing the remaining twenty to thirty percent of blood into the ventricles. This completes ventricular filling (end-diastolic volume, EDV).
3. Ventricular Systole (Isovolumetric Contraction): The impulse spreads through the AV node, bundle branches, and Purkinje fibers, causing ventricular contraction. Ventricular pressure rises rapidly, exceeding atrial pressure and forcing the AV valves closed (producing the S1 sound). However, ventricular pressure is still below the pressure in the pulmonary trunk and aorta, so the semilunar valves remain closed. During this brief phase, all valves are closed, and the ventricles contract isovolumetrically (no blood is ejected, volume remains constant).
4. Ventricular Ejection: Ventricular pressure continues to rise until it exceeds the pressure in the pulmonary trunk and aorta. The semilunar valves are forced open, and blood is ejected into these major arteries. The volume of blood ejected by each ventricle per beat is the stroke volume (SV). The volume remaining in each ventricle after ejection is the end-systolic volume (ESV).
5. Early Diastole (Isovolumetric Relaxation): Ventricular repolarization occurs, and the ventricles begin to relax. Ventricular pressure drops rapidly, falling below the pressure in the pulmonary trunk and aorta. Blood starts to flow back towards the ventricles, closing the semilunar valves (producing the S2 sound). Ventricular pressure is still higher than atrial pressure, so the AV valves remain closed. During this phase, all valves are again closed, and the ventricles relax isovolumetrically.
6. Rapid Ventricular Filling: As ventricular pressure continues to fall below atrial pressure, the AV valves open, and blood rushes rapidly from the atria into the ventricles. This marks the beginning of the next cardiac cycle.
The heart’s performance is quantified by cardiac output (CO), the volume of blood pumped by each ventricle per minute. It is calculated as the product of heart rate (HR, beats per minute) and stroke volume (SV, milliliters per beat): CO = HR x SV. Cardiac output is a critical determinant of tissue perfusion and oxygen delivery. It is tightly regulated by the autonomic nervous system (sympathetic and parasympathetic divisions), hormones (e.g., epinephrine), and local factors (e.g., metabolites) to meet the body’s changing demands, such as during exercise, stress, or rest. The heart itself requires a constant supply of oxygen and nutrients to function. This is provided by the coronary circulation, a network of arteries and veins that branch off the aorta and encircle the heart muscle. Blockage of coronary arteries (e.g., by atherosclerosis) can lead to myocardial ischemia (insufficient oxygen) and potentially a heart attack (myocardial infarction).
The Intricate Network: Tracing the Pathways of Circulation
Blood does not simply slosh around the body randomly. It flows through a highly organized, closed-loop system of blood vessels, creating two distinct but interconnected circuits: the pulmonary circulation and the systemic circulation. This dual-circuit design is essential for efficiently oxygenating blood and delivering it to tissues while keeping oxygenated and deoxygenated blood largely separate.
The pulmonary circulation is the shorter, lower-pressure circuit responsible for transporting blood to the lungs and back. Its primary function is gas exchange: loading oxygen onto hemoglobin and unloading carbon dioxide. Deoxygenated blood, low in oxygen and high in carbon dioxide, is pumped from the right ventricle of the heart into the pulmonary trunk. The pulmonary trunk quickly divides into the right and left pulmonary arteries, which carry the blood to the respective lungs. Within the lungs, these arteries branch extensively, forming smaller arterioles and eventually the vast network of pulmonary capillaries. These capillaries surround the alveoli, the tiny air sacs where gas exchange occurs. The walls of the alveoli and the capillaries are extremely thin (forming the respiratory membrane), allowing oxygen to diffuse from the alveolar air into the blood, binding to hemoglobin in the red blood cells. Simultaneously, carbon dioxide diffuses from the blood into the alveoli to be exhaled. After gas exchange, the blood is now oxygenated. It flows from the pulmonary capillaries into pulmonary venules, which merge to form the four pulmonary veins (two from each lung). These pulmonary veins carry the freshly oxygenated blood back to the left atrium of the heart, completing the pulmonary circuit. Notably, the pulmonary arteries are the only arteries in the body that carry deoxygenated blood, and the pulmonary veins are the only veins that carry oxygenated blood.
The systemic circulation is the much larger, higher-pressure circuit that carries oxygenated blood from the heart to all body tissues (except the lungs) and returns deoxygenated blood back to the heart. Oxygenated blood enters the systemic circuit from the left ventricle, which pumps it forcefully into the aorta, the largest artery in the body. The aorta arches over the heart and descends through the thoracic and abdominal cavities. Along its course, the aorta and its major branches (such as the brachiocephalic trunk, left common carotid artery, left subclavian artery, celiac trunk, superior mesenteric artery, renal arteries, and common iliac arteries) distribute oxygenated blood to the head, neck, upper limbs, thoracic organs, abdominal organs, and lower limbs. These arteries branch repeatedly, forming smaller arteries and then arterioles. Arterioles are the primary resistance vessels in the systemic circulation. Their smooth muscle walls can constrict (vasoconstriction) or relax (vasodilation), dramatically altering blood flow into specific capillary beds to match tissue demands. This regulation is crucial for directing blood flow to active tissues (like muscles during exercise) and regulating blood pressure.
From the arterioles, blood enters the capillaries, the microscopic vessels that form the exchange sites between blood and tissues. Capillaries are composed of a single layer of endothelial cells, often with pores (fenestrations) or intercellular clefts, allowing for the passage of water, small solutes, gases, and sometimes even small proteins. This is where the vital exchange occurs: oxygen and nutrients diffuse out of the blood into the interstitial fluid surrounding cells, and carbon dioxide and other metabolic waste products diffuse from the interstitial fluid into the blood. Some exchange also occurs via filtration (due to hydrostatic pressure) and reabsorption (due to osmotic pressure) across the capillary walls. Capillary beds are intricate networks, often with thoroughfare channels and precapillary sphincters (rings of smooth muscle at the entrance to capillaries) that can regulate blood flow through specific capillaries within a bed.
After passing through the capillaries, blood enters the venules, small vessels that collect blood from the capillary beds. Venules merge to form larger veins. Veins are the capacitance vessels of the circulatory system. They have thinner walls and larger lumens than arteries and are much more distensible. At any given time, about sixty to seventy percent of the body’s total blood volume is within the veins. Veins contain valves, particularly in the limbs, which are folds of the tunica intima (inner layer) that prevent the backflow of blood, ensuring it returns to the heart against gravity. The skeletal muscle pump (contraction of leg muscles compressing veins) and the respiratory pump (pressure changes in the thoracic cavity during breathing) also aid venous return. Deoxygenated blood from the upper body is collected by the superior vena cava, while blood from the lower body is collected by the inferior vena cava. These two large veins empty the deoxygenated blood into the right atrium of the heart, completing the systemic circuit and delivering the blood back to the starting point for the pulmonary circuit.
A crucial aspect of systemic circulation is the hepatic portal system. This is a specialized venous pathway that carries blood from the capillary beds of the digestive organs (stomach, small intestine, large intestine, spleen, pancreas) to the capillary beds within the liver before it returns to the general circulation. Blood leaving these digestive organs, rich in absorbed nutrients but potentially also containing toxins or bacteria, enters the hepatic portal vein. This vein delivers the blood directly to the liver. Within the liver, the blood flows through a second set of capillaries (hepatic sinusoids). This arrangement allows the liver to process, store, modify, or detoxify the absorbed nutrients and other substances before they enter the rest of the systemic circulation via the hepatic veins, which drain into the inferior vena cava. The hepatic portal system is vital for metabolic regulation and detoxification.
Another important specialized circulation is the coronary circulation, mentioned earlier, which supplies the heart muscle itself. Coronary arteries branch from the aorta just above the aortic valve. They run across the surface of the heart and send branches inward to supply the myocardium. Cardiac veins drain blood from the heart muscle; most merge into the coronary sinus, which empties directly into the right atrium. The heart has a very high oxygen extraction rate from its blood supply, making it particularly vulnerable to interruptions in coronary blood flow.
The Symphony of Flow: Hemodynamics and Regulation

